
WORK 4 MODEL

7 € 50

Le Ptéranodon (/təˈrænədɒn/ ; du grec ancien : πτερόν, romanisé : pteron « aile » et ἀνόδων, anodon « édenté ») est un genre de ptérosaure qui comprenait certains des plus grands reptiles volants connus, P. longiceps ayant une envergure de plus de 6 m (20 pi). Ils ont vécu pendant la période géologique de la fin du Crétacé de l’Amérique du Nord dans les actuels Kansas, Nebraska, Wyoming, Dakota du Sud et Alabama. Plus de spécimens fossiles de Ptéranodon ont été trouvés que tout autre ptérosaure, avec environ 1'200 spécimens connus de la science, beaucoup d’entre eux bien conservés avec des crânes presque complets et des squelettes articulés. Il s’agissait d’une partie importante de la communauté animale de la Voie maritime intérieure de l’Ouest.

Lorsque les premiers fossiles de Ptéranodon ont été trouvés, ils ont été attribués à des genres de ptérosaures à dents, tels que Ornithocheirus et Pterodactylus. En 1876, Othniel Charles Marsh l’a reconnu comme un genre à part entière, en notant en particulier son absence totale de dents, ce qui était à l’époque unique parmi les ptérosaures. Au fil des décennies, de multiples espèces ont été attribuées au Pteranodon, bien qu’aujourd’hui, seules 2 soient reconnues : P. longiceps, l’espèce type, et P. sternbergi. Une troisième espèce, P. maiseyi, peut également exister. Certains chercheurs ont suggéré que ces 2 derniers étaient un genre à part entière, Geosternbergia, bien que cela fasse l’objet d’un débat. Un autre genre séparé de Pteranodon, Dawndraco, peut être synonyme de Geosternbergia si ce genre est valide, ou de Pteranodon s’il ne l’est pas.
Le Ptéranodon fait partie de la famille des Pteranodontidae, qui fait partie du clade Pteranodontia, qui comprend également les nyctosauridés. Les ptéranodontes forment un clade plus grand, les Pteranodontoidea, aux côtés des ornithocheiromorphes, et ce clade appartient au sous-ordre des Pterodactyloidea. Bien qu’ils ne soient pas des dinosaures, les ptérosaures tels que Ptéranodon forment un clade étroitement lié aux dinosaures car les 2 appartiennent au clade des Avemetatarsalia.
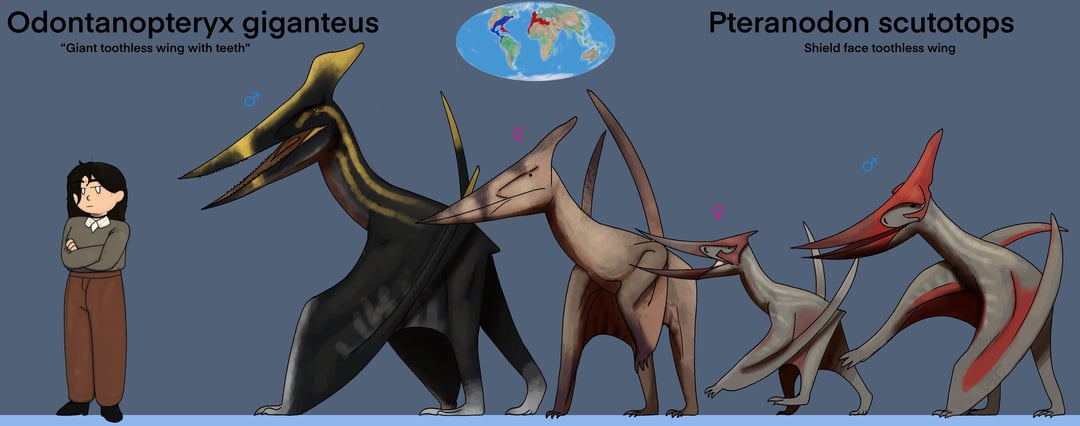
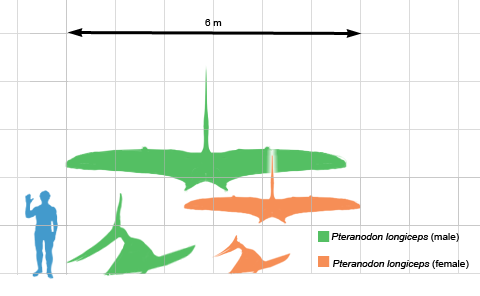
Les ptéranodons mâles et femelles différaient par leur taille et la forme de leur crête. Les mâles ont atteint une envergure de 5,6 à 7,6 m (18 à 25 pi) ; les femelles étaient plus petites, mesurant en moyenne 3,8 m (12 pi). Les crêtes des mâles étaient beaucoup plus grandes que celles des femelles. Chez le P. longiceps, elles étaient longues et balayées vers le dos, tandis que chez le P. sternbergi, elles étaient grandes et droits. Les femelles avaient également un bassin plus large que les mâles.

|
Le Ptéranodon a été le premier ptérosaure trouvé en dehors de l’Europe. Ses fossiles ont été découverts pour la première fois par Othniel Charles Marsh en 1871, dans les dépôts de craie de Smoky Hill du Crétacé supérieur de l’ouest du Kansas. Ces lits de craie se sont déposés au fond de ce qui était autrefois la Voie maritime intérieure de l’Ouest, une grande mer peu profonde au-dessus de ce qui est aujourd’hui la partie médiane du continent nord-américain. Ces premiers spécimens, YPM 1160 et YPM 1161, se composaient d’os partiels des ailes, ainsi que d’une dent du poisson préhistorique Xiphactinus, que Marsh croyait à tort appartenir à ce nouveau ptérosaure (tous les ptérosaures connus jusqu’à ce moment-là avaient des dents). En 1871, Marsh a nommé la découverte Pterodactylus oweni, l’attribuant au genre européen bien connu (mais beaucoup plus petit) Pterodactylus. Marsh a également collecté d’autres os des ailes du grand ptérosaure en 1871. Réalisant que le nom qu’il avait choisi avait déjà été utilisé pour l’espèce européenne de ptérosaure Pterodactylus oweni de Harry Seeley en 1864, Marsh a renommé son ptérosaure géant d’Amérique du Nord Pterodactylus occidentalis, ce qui signifie « doigt de l’aile occidentale », dans sa description de 1872 du nouveau spécimen. Il a nommé 2 espèces supplémentaires, basées sur les différences de taille : Pterodactylus ingens (le plus grand spécimen à ce jour) et Pterodactylus velox (le plus petit).
Entre-temps, le rival de Marsh, Edward Drinker Cope, avait déterré plusieurs spécimens du grand ptérosaure d’Amérique du Nord. Sur la base de ces spécimens, Cope a nommé 2 nouvelles espèces, Ornithochirus umbrosus et Ornithochirus harpyia, dans le but de les attribuer au grand genre européen Ornithocheirus, bien qu’il ait mal orthographié le nom (en oubliant le « e »). L’article de Cope nommant son espèce a été publié en 1872, 5 jours seulement après l’article de Marsh. Cela a donné lieu à une dispute, disputée dans la littérature publiée, pour savoir quels noms avaient la priorité dans ce qui était manifestement la même espèce. Cope concéda en 1875 que les noms de Marsh avaient la priorité sur les siens, mais maintint que Pterodactylus umbrosus était une espèce distincte (mais pas un genre) de toutes celles que Marsh avait nommées auparavant. Une réévaluation par des scientifiques ultérieurs a soutenu le cas de Marsh, réfutant l’affirmation de Cope selon laquelle P. umbrosus représentait une espèce plus grande et distincte.

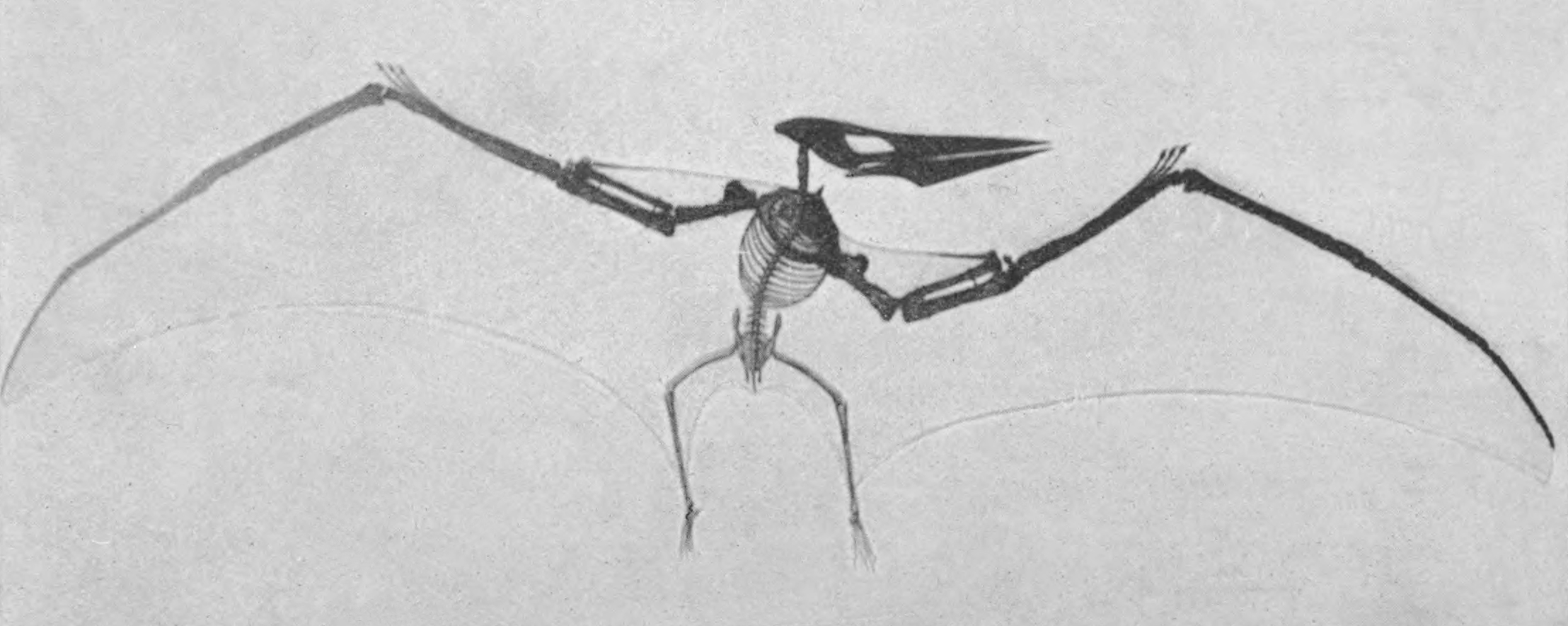
Alors que les premiers os d’ailes de ptéranodon ont été collectés par Marsh et Cope au début des années 1870, le premier crâne de ptéranodon a été trouvé le 2 mai 1876 le long de la rivière Smoky Hill dans le comté de Wallace (aujourd’hui le comté de Logan), au Kansas, aux États-Unis, par Samuel Wendell Williston, un collectionneur de fossiles travaillant pour Marsh. Un deuxième crâne, plus petit, a rapidement été découvert. Ces crânes ont montré que les ptérosaures d’Amérique du Nord étaient différents de toutes les espèces européennes, en ce sens qu’ils n’avaient pas de dents et avaient des crêtes osseuses sur leur crâne. Marsh a reconnu cette différence majeure, décrivant les spécimens comme « se distinguant de tous les genres précédemment connus de l’ordre des Ptérosaures par l’absence totale de dents ». Marsh a reconnu que cette caractéristique justifiait un nouveau genre, et il a inventé le nom de Pteranodon (« aile sans dent ») en 1876. Marsh a reclassifié toutes les espèces nord-américaines précédemment nommées, de Pterodactylus à Pteranodon. Il a considéré que le plus petit crâne appartenait à Pteranodon occidentalis, en fonction de sa taille. Marsh a classé le plus grand crâne, YPM 1117, dans la nouvelle espèce Pteranodon longiceps, qu’il pensait être une espèce de taille moyenne entre le petit P. occidentalis et le grand P. ingens. Marsh a également nommé plusieurs espèces supplémentaires : Pteranodon comptus et Pteranodon nanus ont été nommés d’après des squelettes fragmentaires de petits individus, tandis que Pteranodon gracilis était basé sur un os de l’aile qu’il a confondu avec un os pelvien. Il s’est vite rendu compte de son erreur et a reclassé ce spécimen dans un genre distinct, qu’il a nommé Nyctosaurus. Le P. nanus a également été reconnu plus tard comme un spécimen de Nyctosaurus.
 En 1892, Samuel Williston a examiné la question de la classification des ptéranodons. Il remarqua qu’en 1871, Seeley avait mentionné l’existence d’un ensemble partiel de mâchoires de ptérosaure édentées provenant des Cambridge Greensand d’Angleterre, qu’il nomma Ornithostoma. Parce que la principale caractéristique que Marsh avait utilisée pour séparer le ptéranodon des autres ptérosaures était son manque de dents, Williston a conclu que « Ornithostoma » doit être considéré comme le synonyme principal de Ptéranodon. Cependant, en 1901, Pleininger a souligné que « Ornithostoma » n’avait jamais été scientifiquement décrit ni même attribué un nom d’espèce jusqu’aux travaux de Williston, et avait donc été un nomen nudum et ne pouvait pas battre Pteranodon pour la priorité de nommage. Williston a accepté cette conclusion et est retourné à l’appel du genre Pteranodon. Cependant, Williston et Pleininger avaient tous 2 tort, car ils n’ont pas remarqué le fait qu’en 1891, Seeley lui-même avait finalement décrit et correctement nommé Ornithostoma, l’attribuant à l’espèce O. sedgwicki. Dans les années 2010, d’autres recherches sur l’identité d’Ornithostoma ont montré qu’il ne s’agissait probablement pas d’un ptéranodon ou même d’un proche parent, mais qu’il pourrait en fait s’agir d’un azhdarchoïde, un autre type de ptérosaure édenté.
En 1892, Samuel Williston a examiné la question de la classification des ptéranodons. Il remarqua qu’en 1871, Seeley avait mentionné l’existence d’un ensemble partiel de mâchoires de ptérosaure édentées provenant des Cambridge Greensand d’Angleterre, qu’il nomma Ornithostoma. Parce que la principale caractéristique que Marsh avait utilisée pour séparer le ptéranodon des autres ptérosaures était son manque de dents, Williston a conclu que « Ornithostoma » doit être considéré comme le synonyme principal de Ptéranodon. Cependant, en 1901, Pleininger a souligné que « Ornithostoma » n’avait jamais été scientifiquement décrit ni même attribué un nom d’espèce jusqu’aux travaux de Williston, et avait donc été un nomen nudum et ne pouvait pas battre Pteranodon pour la priorité de nommage. Williston a accepté cette conclusion et est retourné à l’appel du genre Pteranodon. Cependant, Williston et Pleininger avaient tous 2 tort, car ils n’ont pas remarqué le fait qu’en 1891, Seeley lui-même avait finalement décrit et correctement nommé Ornithostoma, l’attribuant à l’espèce O. sedgwicki. Dans les années 2010, d’autres recherches sur l’identité d’Ornithostoma ont montré qu’il ne s’agissait probablement pas d’un ptéranodon ou même d’un proche parent, mais qu’il pourrait en fait s’agir d’un azhdarchoïde, un autre type de ptérosaure édenté.
 Williston a également été le premier scientifique à évaluer de manière critique toutes les espèces de ptéranodons classées par Cope et Marsh. Il était d’accord avec la plupart des classifications de Marsh, à quelques exceptions près. Premièrement, il ne croyait pas que le P. ingens et le P. umbrosus pouvaient être considérés comme des synonymes, ce que même Cope en était venu à croire. Il considérait que le P. velox et le P. longiceps étaient douteux ; la première était basée sur des fragments non diagnostiques, et la seconde, bien que connue à partir d’un crâne complet, appartenait probablement à l’une des autres espèces précédemment nommées. En 1903, Williston réexamina la question de la classification des ptéranodons et révisa sa conclusion antérieure selon laquelle il y avait 7 espèces à seulement 3. Il considérait le P. comptus et le P. nanus comme des spécimens de Nyctosaurus, et a divisé les autres en petites espèces (P. velox), moyennes (P. occidentalis) et grandes (P. ingens), en se basant principalement sur la forme de leurs os du bras. Il pensait que le P. longiceps, le seul connu d’un crâne, pourrait être un synonyme du P. velox ou du P. occidentalis, en fonction de sa taille.
Williston a également été le premier scientifique à évaluer de manière critique toutes les espèces de ptéranodons classées par Cope et Marsh. Il était d’accord avec la plupart des classifications de Marsh, à quelques exceptions près. Premièrement, il ne croyait pas que le P. ingens et le P. umbrosus pouvaient être considérés comme des synonymes, ce que même Cope en était venu à croire. Il considérait que le P. velox et le P. longiceps étaient douteux ; la première était basée sur des fragments non diagnostiques, et la seconde, bien que connue à partir d’un crâne complet, appartenait probablement à l’une des autres espèces précédemment nommées. En 1903, Williston réexamina la question de la classification des ptéranodons et révisa sa conclusion antérieure selon laquelle il y avait 7 espèces à seulement 3. Il considérait le P. comptus et le P. nanus comme des spécimens de Nyctosaurus, et a divisé les autres en petites espèces (P. velox), moyennes (P. occidentalis) et grandes (P. ingens), en se basant principalement sur la forme de leurs os du bras. Il pensait que le P. longiceps, le seul connu d’un crâne, pourrait être un synonyme du P. velox ou du P. occidentalis, en fonction de sa taille.
 En 1910, Eaton est devenu le premier scientifique à publier une description plus détaillée de l’ensemble du squelette du ptéranodon, tel qu’il était connu à l’époque. Il a utilisé ses résultats pour réviser la classification du genre une fois de plus sur la base d’une meilleure compréhension des différences dans l’anatomie des ptéranodontes. Eaton a mené des expériences en utilisant des modèles d’os en argile pour aider à déterminer les effets de l’écrasement et de l’aplatissement sur les formes des os du bras que Williston avait utilisés dans sa propre classification. Eaton a découvert que la plupart des différences dans les formes des os pouvaient être facilement expliquées par les pressions de la fossilisation, et a conclu qu’aucun squelette de ptéranodon n’avait de différences significatives les uns par rapport aux autres en dehors de leur taille. Par conséquent, Eaton a dû décider de son système de classification en se basant uniquement sur les différences dans les crânes, qu’il a attribués aux espèces tout comme Marsh l’a fait, en fonction de leur taille. En fin de compte, Eaton n’a reconnu que 3 espèces valides : P. occidentalis, P. ingens et P. longiceps.
En 1910, Eaton est devenu le premier scientifique à publier une description plus détaillée de l’ensemble du squelette du ptéranodon, tel qu’il était connu à l’époque. Il a utilisé ses résultats pour réviser la classification du genre une fois de plus sur la base d’une meilleure compréhension des différences dans l’anatomie des ptéranodontes. Eaton a mené des expériences en utilisant des modèles d’os en argile pour aider à déterminer les effets de l’écrasement et de l’aplatissement sur les formes des os du bras que Williston avait utilisés dans sa propre classification. Eaton a découvert que la plupart des différences dans les formes des os pouvaient être facilement expliquées par les pressions de la fossilisation, et a conclu qu’aucun squelette de ptéranodon n’avait de différences significatives les uns par rapport aux autres en dehors de leur taille. Par conséquent, Eaton a dû décider de son système de classification en se basant uniquement sur les différences dans les crânes, qu’il a attribués aux espèces tout comme Marsh l’a fait, en fonction de leur taille. En fin de compte, Eaton n’a reconnu que 3 espèces valides : P. occidentalis, P. ingens et P. longiceps.
 La découverte de spécimens à crêtes dressées, classés par Harksen en 1966 comme la nouvelle espèce de Pteranodon sternbergi, a encore compliqué la situation ce qui a entraîné une autre révision du genre par Halsey W. Miller en 1972. Parce qu’il était impossible de déterminer la forme de la crête de toutes les espèces sur la base de squelettes sans tête, Miller a conclu que toutes les espèces de Ptéranodon, à l’exception des 2 basées sur les crânes (P. longiceps et P. sternbergi) doivent être considérées comme nomena dubia et abandonnées. Le crâne qu’Eaton pensait appartenir à P. ingens a été placé dans la nouvelle espèce Pteranodon marshi, et le crâne qu’Eaton a attribué à P. occidentalis a été renommé Pteranodon eatoni. Miller a également reconnu une autre espèce basée sur un crâne avec une crête similaire à celle du P. sternbergi ; Miller a nommé ce Ptéranodon walkeri. Pour aider à mettre de l’ordre dans cet enchevêtrement de noms, Miller a créé 3 sous-genres. Le P. marshi et le P. longiceps ont été placés dans le sous-genre Longicepia, bien que celui-ci ait ensuite été changé en Ptéranodon en raison des règles de priorité. Le P. sternbergi et le P. walkeri, l’espèce à crête dressée, ont reçu le sous-genre Sternbergia, qui a ensuite été changé en Geosternbergia parce que Sternbergia était préoccupé. Enfin, Miller a nommé le sous-genre Occidentalia d’après le P. eatoni, le crâne anciennement associé au P. occidentalis. Miller a encore élargi le concept de ptéranodon pour inclure Nyctosaurus en tant que quatrième sous-genre. Miller considérait qu’il s’agissait d’une progression évolutive, le Nyctosaurus primitif, à l’époque considéré comme sans crête, donnant naissance à des Occidentalia à petite crête, qui à leur tour ont donné naissance à des Ptéranodon à longue crête, conduisant finalement à des Geosternbergia à grande crête. Cependant, Miller a commis plusieurs erreurs dans son étude concernant les spécimens que Marsh avait attribués à quelle espèce, et la plupart des scientifiques ont ignoré son travail sur le sujet dans leurs recherches ultérieures. En 1984, Robert Milton Schoch a publié une autre révision qui revenait essentiellement au schéma de classification original de Marsh, notamment en faisant couler le P. longiceps comme synonyme du P. ingens.
La découverte de spécimens à crêtes dressées, classés par Harksen en 1966 comme la nouvelle espèce de Pteranodon sternbergi, a encore compliqué la situation ce qui a entraîné une autre révision du genre par Halsey W. Miller en 1972. Parce qu’il était impossible de déterminer la forme de la crête de toutes les espèces sur la base de squelettes sans tête, Miller a conclu que toutes les espèces de Ptéranodon, à l’exception des 2 basées sur les crânes (P. longiceps et P. sternbergi) doivent être considérées comme nomena dubia et abandonnées. Le crâne qu’Eaton pensait appartenir à P. ingens a été placé dans la nouvelle espèce Pteranodon marshi, et le crâne qu’Eaton a attribué à P. occidentalis a été renommé Pteranodon eatoni. Miller a également reconnu une autre espèce basée sur un crâne avec une crête similaire à celle du P. sternbergi ; Miller a nommé ce Ptéranodon walkeri. Pour aider à mettre de l’ordre dans cet enchevêtrement de noms, Miller a créé 3 sous-genres. Le P. marshi et le P. longiceps ont été placés dans le sous-genre Longicepia, bien que celui-ci ait ensuite été changé en Ptéranodon en raison des règles de priorité. Le P. sternbergi et le P. walkeri, l’espèce à crête dressée, ont reçu le sous-genre Sternbergia, qui a ensuite été changé en Geosternbergia parce que Sternbergia était préoccupé. Enfin, Miller a nommé le sous-genre Occidentalia d’après le P. eatoni, le crâne anciennement associé au P. occidentalis. Miller a encore élargi le concept de ptéranodon pour inclure Nyctosaurus en tant que quatrième sous-genre. Miller considérait qu’il s’agissait d’une progression évolutive, le Nyctosaurus primitif, à l’époque considéré comme sans crête, donnant naissance à des Occidentalia à petite crête, qui à leur tour ont donné naissance à des Ptéranodon à longue crête, conduisant finalement à des Geosternbergia à grande crête. Cependant, Miller a commis plusieurs erreurs dans son étude concernant les spécimens que Marsh avait attribués à quelle espèce, et la plupart des scientifiques ont ignoré son travail sur le sujet dans leurs recherches ultérieures. En 1984, Robert Milton Schoch a publié une autre révision qui revenait essentiellement au schéma de classification original de Marsh, notamment en faisant couler le P. longiceps comme synonyme du P. ingens.
À la fin des années 1980 et au début des années 1990, S. Christopher Bennett a publié plusieurs articles importants sur l’anatomie, la taxonomie et l’histoire de la vie du ptéranodon. En 1992, il a publié un article sur le dimorphisme sexuel et son rôle dans la variation individuelle parmi les fossiles de ptéranodon, une suite d’un article de 1987 qu’il a écrit sur le même sujet. Dans l’article de 1992, il ne faisait référence qu’à 2 espèces, le P. longiceps et le P. sternbergi. 2 ans plus tard, il publia un article révisant entièrement sa taxonomie, dans lequel il concluait que seules le P. longiceps et le P. sternbergi étaient des espèces valides. Le P. marshi et le P. walkeri étaient considérés comme des synonymes juniors du P. longiceps, et le P. eatoni comme un synonyme junior du P. stenbergi. Le reste a été soit rendu nomina dubia, soit placé dans Nyctosaurus.
|
Les ptéranodons mâles adultes étaient parmi les plus grands ptérosaures et étaient les plus grands animaux volants connus jusqu’à la fin du XXe siècle, lorsque les ptérosaures géants azhdarchidés ont été découverts. L’envergure d’un ptéranodon mâle adulte moyen était de 5,6 m (18 pi). Les femelles adultes étaient beaucoup plus petites, mesurant en moyenne 3,8 m (12 pi) d’envergure. On estime qu’un grand spécimen de Pteranodon longiceps, USNM 50130, a une envergure de 6,25 à 6,5 m (20,5 à 21,3 pi), une longueur de corps de 2,6 m (8,5 pi) et une masse corporelle de 50 kg (110 lb). Des spécimens encore plus grands avaient une envergure de 7,25 à 7,6 m (23,8 à 24,9 pieds). Mis à part la taille, les femelles se distinguaient par leurs crêtes cérébrales courtes et arrondies et leurs larges canaux pelviens, tandis que les mâles avaient des hanches étroites et de très grandes crêtes cérébrales, remplissant probablement une fonction de parade.
Les méthodes utilisées pour estimer la masse des grands spécimens de ptéranodon mâles (ceux dont l’envergure est d’environ 7 mètres) ont été notoirement peu fiables, produisant un large éventail d’estimations. Dans une revue des estimations de la taille des ptérosaures publiée en 2010, Mark Witton et Michael Habib ont fait valoir que la plus grande estimation de 93 kg (205 lb) est beaucoup trop élevée et qu’une limite supérieure de 20 à 35 kg (44 à 77 lb) est plus réaliste. Witton et Habib ont considéré que les méthodes utilisées par les chercheurs qui ont obtenu des estimations de masse plus petites étaient tout aussi erronées. La plupart ont été produits en mettant à l’échelle des animaux modernes tels que les chauves-souris et les oiseaux jusqu’à la taille d’un ptéranodon, malgré le fait que les ptérosaures ont des proportions corporelles et une anatomie des tissus mous très différentes de celles de tout animal vivant.

|
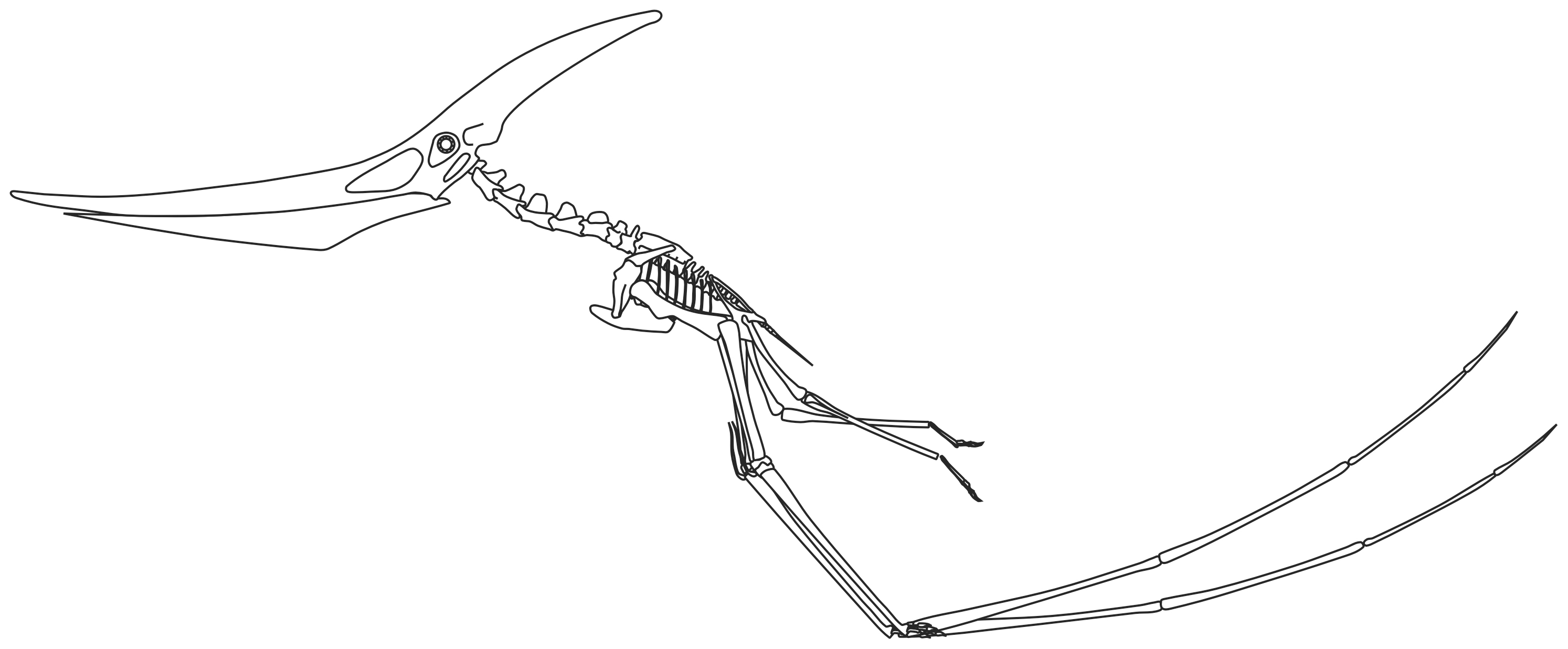
Contrairement aux ptérosaures antérieurs, tels que Rhamphorhynchus et Pterodactylus, le ptéranodon avait des becs édentés, similaires à ceux des oiseaux. Les becs des ptéranodons étaient constitués de marges solides et osseuses qui faisaient saillie à partir de la base des mâchoires. Les becs étaient longs, minces et se terminaient par des pointes fines et acérées. La mâchoire supérieure, qui était plus longue que la mâchoire inférieure, était courbée vers le haut ; alors que cela n’a normalement été attribué qu’au bec courbé vers le haut, un spécimen (UALVP 24238) a une courbure correspondant à l’élargissement du bec vers l’extrémité. Bien que l’extrémité du bec ne soit pas connue chez ce spécimen, le niveau de courbure suggère qu’il aurait été extrêmement long. La forme unique du bec de ce spécimen a conduit Alexander Kellner à l’attribuer à un genre distinct, Dawndraco, en 2010.
La caractéristique la plus distinctive du ptéranodon est sa crête crânienne. Ces crêtes étaient constituées d’os du crâne (frontaux) faisant saillie vers le haut et vers l’arrière du crâne. La taille et la forme de ces crêtes variaient en raison d’un certain nombre de facteurs, notamment l’âge, le sexe et l’espèce. Le Pteranodon sternbergi mâle, la plus ancienne des 2 espèces décrites à ce jour, avait une crête plus verticale avec une large projection vers l’avant, tandis que leurs descendants, le Pteranodon longiceps, ont développé une crête plus étroite et plus projetée vers l’arrière. Les femelles des 2 espèces étaient plus petites et portaient de petites crêtes arrondies. Les écussons étaient probablement principalement des structures d’affichage, bien qu’ils aient pu avoir d’autres fonctions.

|
Les épines neurales des vertèbres du ptéranodon étaient étroites. Comme beaucoup de ptérosaures et d’oiseaux, il possédait un notarium, une masse soudée comprenant les 6 premières vertèbres dorsales. De même, les premières côtes ont été fusionnées. Les os pelviens ont été fusionnés au synsacrum, une masse de vertèbres qui comprenait au moins 2 vertèbres dorsales, les vertèbres sacrées et la première vertèbre caudale. Les sacrés étaient renforcés par des ligaments osseux. Au-delà du synsacrum, la queue était relativement courte et les dernières vertèbres étaient fusionnées en une tige osseuse. La longueur totale de la queue était d’environ 3,5% de la longueur de l’envergure des ailes, ou jusqu’à 25 cm (9,8 po) chez les plus grands mâles.
Les omoplates des ptéranodons étaient orientées de telle manière que chacune d’entre elles s’appuie sur l’autre, en raison de leur fusion avec les coracoïdes, offrant une intégrité accrue pendant le vol. Les humérus étaient extrêmement robustes, avec de grandes crêtes delto-pectorales incurvées. Le radius et le cubitus étaient tout aussi robustes. Les 3 premiers métacarpiens étaient très minces et leurs doigts respectifs arboraient des ongulés courts et incurvés (griffes). Les pattes arrière du ptéranodon avaient 4 métatarsiens, qui étaient terminés par des griffes moins incurvées.
|
La forme de l’aile du ptéranodon suggère qu’il aurait volé un peu comme un albatros des temps modernes. Ceci est basé sur le fait que le ptéranodon avait un rapport d’aspect élevé (envergure par rapport à la longueur de la corde) similaire à celui de l’albatros - 9:1 pour le ptéranodon, contre 8:1 pour un albatros. Les albatros passent de longues périodes de temps à pêcher en mer et utilisent un modèle de vol appelé « soaring dynamique » qui exploite le gradient vertical de la vitesse du vent près de la surface de l’océan pour parcourir de longues distances sans battre des ailes et sans l’aide de thermiques (qui ne se produisent pas au-dessus de l’océan ouvert de la même manière qu’ils le font au-dessus de la terre). Bien que la majeure partie du vol d’un ptéranodon ait dépendu du plané, comme les oiseaux de mer à longues ailes, il a probablement nécessité une rafale active et rapide occasionnelle de battements, et des études de la charge alaire des ptéranodon (la force des ailes par rapport au poids du corps) indiquent qu’ils étaient capables de voler en battant des ailes substantiellement, contrairement à certaines suggestions antérieures selon lesquelles ils étaient si grands qu’ils ne pouvaient que planer. Cependant, une étude plus récente suggère qu’il s’appuyait sur le soaring thermique, contrairement aux oiseaux de mer modernes, mais un peu comme les volants continentaux modernes et les Pelagornis éteints.
Comme d’autres ptérosaures, le Ptéranodon a probablement décollé d’une position debout et quadrupède. En utilisant leurs longs membres antérieurs comme levier, ils se seraient élancés dans les airs d’un bond rapide. Presque toute l’énergie aurait été générée par les membres antérieurs. La course ascendante des ailes se serait produite lorsque l’animal aurait dégagé le sol, suivie d’une descente rapide pour générer une portance supplémentaire et terminer le lancement dans les airs. Il est possible que le Ptéranodon ait pu réaliser cela depuis l’eau, ainsi que sur terre, ce qui a été spéculé pour divers autres tels que l’Anhanguera, parent éloigné.

Historiquement, la locomotion terrestre chez les ptéranodons, comme chez les ptérosaures en général, a fait l’objet de débats, principalement de savoir s’ils étaient bipèdes ou quadrupèdes. Le premier modèle de locomotion des ptéranodons, proposé par Cherrie D. Bramwell et G. R. Whitfield, suggérait qu’ils étaient totalement incapables de marcher ou de se tenir debout. Au lieu de cela, ils ont suggéré qu’il se déplaçait sur la terre ferme en se poussant, et qu’il avait décollé en se perchant sur les falaises et en laissant le vent l’emporter. Les travaux ultérieurs ont largement tourné autour de méthodes de locomotion plus conventionnelles, telles que la bipédie et divers types de quadrupède. En 2004, Sankar Chatterjee et R. J. Templin ont proposé un système double, dans lequel les ptérosaures marchaient en quadrupède la plupart du temps, mais ont opté pour un décollage bipède. Ce dernier cas, cependant, est peu probable. Les traces suggèrent que les ptérosaures comme le Ptéranodon étaient quadrupèdes.
On sait que le régime alimentaire du ptéranodon comprenait du poisson ; des arêtes de poisson fossilisées ont été trouvées dans la région de l’estomac d’un ptéranodon, et un bolus de poisson fossilisé a été trouvé entre les mâchoires d’un autre ptéranodon, le spécimen AMNH 5098. De nombreux autres spécimens conservent également des fragments d’écailles de poisson et de vertèbres près du torse, indiquant que les poissons constituaient la majorité du régime alimentaire de Ptéranodon (bien qu’ils aient peut-être aussi pris des invertébrés).

Traditionnellement, la plupart des chercheurs ont suggéré que le Pteranodon aurait pris des poissons en plongeant leur bec dans l’eau alors qu’ils volaient à basse altitude. Cependant, cela était probablement basé sur l’hypothèse que les animaux ne pouvaient pas décoller de la surface de l’eau. Il est plus probable que le ptéranodon aurait pu décoller de l’eau et aurait plongé pour attraper des poissons en nageant plutôt qu’en volant. Même une petite femelle ptéranodon aurait pu atteindre une profondeur d’au moins 80 centimètres (31 po) avec son long bec et son cou tout en flottant à la surface, et ils auraient pu atteindre des profondeurs encore plus grandes en plongeant dans l’eau depuis les airs comme certains oiseaux de mer modernes à longues ailes. En 1994, Bennett a noté que la tête, le cou et les épaules du ptéranodon étaient aussi lourds que ceux des oiseaux plongeurs, et a suggéré qu’ils pouvaient plonger en repliant leurs ailes comme le fou de Bassan moderne.
Le ptéranodon était remarquable pour sa crête crânienne, bien que la fonction de cette crête ait fait l’objet d’un débat. Cependant, la plupart des explications se sont concentrées sur la crête en forme de lame et pointue vers l’arrière du mâle P. longiceps et ont ignoré la large gamme de variations selon l’âge et le sexe. Le fait que les crêtes varient autant exclut la plupart des fonctions pratiques autres que l’utilisation dans les parades nuptiales. Par conséquent, l’affichage était probablement la fonction principale de la crête, et toutes les autres fonctions étaient secondaires.

Les interprétations scientifiques de la fonction de la crête ont commencé en 1910, lorsque George Francis Eaton a proposé 2 possibilités : un contrepoids aérodynamique et un point d’attache musculaire. Il a suggéré que la crête aurait pu ancrer de grands et longs muscles de la mâchoire, mais a admis que cette fonction seule ne pouvait pas expliquer la grande taille de certaines crêtes. Bennett (1992) était d’accord avec l’évaluation d’Eaton elle-même selon laquelle la crête était trop grande et trop variable pour avoir été un site d’attache musculaire. Eaton avait suggéré qu’une fonction secondaire de la crête aurait pu être un contrepoids contre le long bec, réduisant ainsi le besoin de muscles lourds du cou pour contrôler l’orientation de la tête. Des essais en soufflerie ont montré que la crête fonctionnait comme un contrepoids efficace dans une certaine mesure, mais Bennett a noté que, encore une fois, l’hypothèse se concentre uniquement sur les longues crêtes de P. longiceps mâles, et non sur les crêtes plus grandes de P. sternbergi et les très petites crêtes qui existaient parmi les femelles. Bennett a constaté que les crêtes des femelles n’avaient pas d’effet compensatoire, et que les crêtes des P. sternbergi mâles auraient, en elles-mêmes, un effet négatif sur l’équilibre de la tête. En fait, le mouvement latéral des crêtes aurait nécessité plus, et non moins, de musculature du cou pour contrôler l’équilibre.

En 1943, Dominik von Kripp a suggéré que la crête aurait pu servir de gouvernail, une idée adoptée par plusieurs chercheurs ultérieurs. Un chercheur, Ross S. Stein, a même suggéré que la crête aurait pu soutenir une membrane de peau reliant la crête pointant vers l’arrière au cou et au dos, augmentant ainsi sa surface et son efficacité en tant que gouvernail. L’hypothèse du gouvernail, encore une fois, ne prend pas en compte les femelles ni les P. sternbergi, qui avaient une crête pointant vers le haut et non vers l’arrière. Bennett a également constaté que, même en tant que gouvernail, la crête ne fournirait pas autant de force directionnelle que la simple manœuvre des ailes. La suggestion que la crête était un frein à air et que les animaux tournaient la tête sur le côté afin de ralentir souffre d’un problème similaire. De plus, les hypothèses de la gouverne de direction et des aérofreins n’expliquent pas pourquoi il existe une telle variation de la taille de la crête, même chez les adultes.
Alexander Kellner a suggéré que les grandes crêtes du ptérosaure Tapejara, ainsi que d’autres espèces, pourraient être utilisées pour l’échange de chaleur, permettant à ces ptérosaures d’absorber ou d’évacuer la chaleur et de réguler la température corporelle, ce qui expliquerait également la corrélation entre la taille de la crête et la taille du corps. Cependant, il n’y a aucune preuve de vaisseaux sanguins supplémentaires dans la crête à cette fin, et les grandes ailes membraneuses remplies de vaisseaux sanguins auraient servi cet objectif beaucoup plus efficacement.
Avec ces hypothèses exclues, l’hypothèse la mieux étayée pour la fonction de la crête semble être une manifestation sexuelle. Cela correspond à la variation de taille observée chez les spécimens fossiles, où les femelles et les juvéniles ont de petites crêtes et les mâles de grandes crêtes élaborées et variables.

Les spécimens adultes de Ptéranodon peuvent être divisés en 2 classes de taille distinctes, petite et grande, la grande classe étant environ une fois et demie plus grande que la petite classe, et la petite classe étant 2 fois plus commune que la grande classe. Les 2 classes de taille vivaient côte à côte, et bien que les chercheurs aient précédemment suggéré qu’elles représentaient des espèces différentes, Christopher Bennett a montré que les différences entre elles sont cohérentes avec le concept selon lequel elles représentent des femelles et des mâles, et que les espèces de ptéranodon étaient sexuellement dimorphes. Les crânes de la classe de taille supérieure conservent de grandes crêtes pointant vers le haut et vers l’arrière, tandis que les crêtes de la classe de taille plus petite sont petites et triangulaires. Certains crânes plus grands montrent également des signes d’une deuxième crête qui s’étendait longue et basse, vers l’extrémité du bec, ce qui n’est pas le cas chez les petits spécimens.

Le sexe des différentes classes de taille a été déterminé, non pas à partir des crânes, mais à partir des os pelviens. Contrairement à ce que l’on pourrait attendre, la classe de taille plus petite avait des os pelviens disproportionnellement grands et larges. Bennett a interprété cela comme indiquant un canal de naissance plus spacieux, par lequel les œufs passeraient. Il a conclu que la classe de petite taille avec de petites crêtes triangulaires représente les femelles, et que les spécimens plus grands et à grande crête représentent les mâles. La taille globale et la taille de la crête correspondent également à l’âge. Les spécimens immatures sont connus à la fois chez les femelles et les mâles, et les mâles immatures ont souvent de petites crêtes similaires à celles des femelles adultes. Par conséquent, il semble que les grandes crêtes ne se soient développées chez les mâles que lorsqu’ils ont atteint leur grande taille adulte, ce qui rend le sexe des spécimens immatures difficile à établir à partir de restes partiels.

Le fait que les femelles semblent avoir été 2 fois plus nombreuses que les mâles suggère que, comme pour les animaux modernes présentant un dimorphisme sexuel lié à la taille, tels que les lions de mer et autres pinnipèdes, le ptéranodon aurait pu être polygyne, avec quelques mâles en compétition pour l’association avec des groupes composés d’un grand nombre de femelles. À l’instar des pinnipèdes modernes, les ptéranodons ont peut-être rivalisé pour établir leur territoire sur des roqueries rocheuses au large, les mâles les plus grands et les plus huppés gagnant le plus de territoire et ayant plus de succès en s’accouplant avec les femelles. Les crêtes des ptéranodons mâles n’auraient pas été utilisées en compétition, mais plutôt comme « symboles visuels de dominance », les rituels de parade remplaçant la compétition physique avec d’autres mâles. Si cette hypothèse est correcte, il est également probable que le Ptéranodon mâle n’a joué que peu ou pas de rôle dans l’élevage des jeunes ; un tel comportement ne se trouve pas chez les mâles des animaux polygynes modernes qui engendrent plusieurs descendants en même temps.
 et d’autres voies maritimes avoisinantes") Des spécimens attribués au Pteranodon ont été trouvés dans les dépôts de craie de Smoky Hill de la formation de Niobrara et dans les dépôts légèrement plus jeunes de Sharon Springs de la formation de schiste de Pierre. Du vivant du Ptéranodon, cette zone était recouverte d’une grande mer intérieure, connue sous le nom de Voie maritime intérieure de l’Ouest. Célèbres pour leurs fossiles collectés depuis 1870, ces formations s’étendent d’aussi loin au sud que le Kansas aux États-Unis jusqu’au Manitoba au Canada. Cependant, des spécimens de ptéranodon (ou tout autre spécimen de ptérosaure) n’ont été trouvés que dans la moitié sud de la formation, au Kansas, au Wyoming et au Dakota du Sud. Malgré le fait que de nombreux fossiles aient été trouvés dans les parties contemporaines de la formation au Canada, aucun spécimen de ptérosaure n’y a jamais été trouvé. Cela suggère fortement que l’aire de répartition géographique naturelle du ptéranodon ne couvrait que la partie sud de la Niobrara, et que son habitat ne s’étendait pas plus au nord que le Dakota du Sud.
Des spécimens attribués au Pteranodon ont été trouvés dans les dépôts de craie de Smoky Hill de la formation de Niobrara et dans les dépôts légèrement plus jeunes de Sharon Springs de la formation de schiste de Pierre. Du vivant du Ptéranodon, cette zone était recouverte d’une grande mer intérieure, connue sous le nom de Voie maritime intérieure de l’Ouest. Célèbres pour leurs fossiles collectés depuis 1870, ces formations s’étendent d’aussi loin au sud que le Kansas aux États-Unis jusqu’au Manitoba au Canada. Cependant, des spécimens de ptéranodon (ou tout autre spécimen de ptérosaure) n’ont été trouvés que dans la moitié sud de la formation, au Kansas, au Wyoming et au Dakota du Sud. Malgré le fait que de nombreux fossiles aient été trouvés dans les parties contemporaines de la formation au Canada, aucun spécimen de ptérosaure n’y a jamais été trouvé. Cela suggère fortement que l’aire de répartition géographique naturelle du ptéranodon ne couvrait que la partie sud de la Niobrara, et que son habitat ne s’étendait pas plus au nord que le Dakota du Sud.
Des fossiles très fragmentaires appartenant à des ptérosaures ptéranodontiens, et peut-être au Ptéranodon lui-même, ont également été trouvés sur la côte du golfe du Mexique et la côte est des États-Unis. Par exemple, certains fragments d’os de la formation de Mooreville en Alabama et de la formation de Merchantville au Delaware peuvent provenir du Pteranodon, bien qu’ils soient trop incomplets pour permettre une identification définitive. Certains restes du Japon ont également été provisoirement attribués au Ptéranodon, mais leur distance par rapport à son habitat connu de la Voie maritime intérieure occidentale rend cette identification peu probable.
 Le Pteranodon longiceps aurait partagé le ciel avec le ptérosaure à crête géante Nyctosaurus. Comparé au P. longiceps, qui était une espèce très commune, le Nyctosaurus était rare, ne représentant que 3% des fossiles de ptérosaures de la formation. L’oiseau à dents précoces, l'Ichthyornis, était également moins commun. Sous la surface, la mer était principalement peuplée d’invertébrés tels que les ammonites et les calmars. La vie vertébrée, en dehors des poissons basaux, comprenait des tortues de mer, telles que les Toxochelys, les plésiosaures Elasmosaurus et les Styxosaurus, et l’oiseau plongeur incapable de voler, le Parahesperornis. Les mosasaures étaient les reptiles marins les plus courants, avec des genres comprenant les Clidastes, les Mosasaurus et les Tylosaurus. Au moins certains de ces reptiles marins sont connus pour s’être nourris de ptéranodon. Barnum Brown, en 1904, a signalé que le contenu de l’estomac d’un plésiosaure contenait des os de ptérodactyle, très probablement de Ptéranodon. Des fossiles de dinosaures terrestres ont également été trouvés dans la craie du Niobrara, ce qui suggère que les animaux morts sur le rivage ont dû être emportés par la mer (un spécimen d’hadrosaure semble avoir été charognardé par un requin).
Le Pteranodon longiceps aurait partagé le ciel avec le ptérosaure à crête géante Nyctosaurus. Comparé au P. longiceps, qui était une espèce très commune, le Nyctosaurus était rare, ne représentant que 3% des fossiles de ptérosaures de la formation. L’oiseau à dents précoces, l'Ichthyornis, était également moins commun. Sous la surface, la mer était principalement peuplée d’invertébrés tels que les ammonites et les calmars. La vie vertébrée, en dehors des poissons basaux, comprenait des tortues de mer, telles que les Toxochelys, les plésiosaures Elasmosaurus et les Styxosaurus, et l’oiseau plongeur incapable de voler, le Parahesperornis. Les mosasaures étaient les reptiles marins les plus courants, avec des genres comprenant les Clidastes, les Mosasaurus et les Tylosaurus. Au moins certains de ces reptiles marins sont connus pour s’être nourris de ptéranodon. Barnum Brown, en 1904, a signalé que le contenu de l’estomac d’un plésiosaure contenait des os de ptérodactyle, très probablement de Ptéranodon. Des fossiles de dinosaures terrestres ont également été trouvés dans la craie du Niobrara, ce qui suggère que les animaux morts sur le rivage ont dû être emportés par la mer (un spécimen d’hadrosaure semble avoir été charognardé par un requin).
Il est probable que, comme chez d’autres animaux polygynes (chez lesquels les mâles sont en compétition pour s’associer aux harems de femelles), les ptéranodons vivaient principalement sur des roqueries au large, où ils pouvaient nicher à l’abri des prédateurs terrestres et se nourrir loin du rivage ; la plupart des fossiles de ptéranodon se trouvent dans des endroits qui, à l’époque, se trouvaient à des centaines de kilomètres de la côte.

Les fossiles de ptéranodon proviennent principalement de la formation de Niobrara, dans le centre des États-Unis. Au sens large, le Ptéranodon a existé pendant plus de 4 millions d’années, au cours de la phase santonienne de la période du Crétacé. Le genre est présent dans la plupart des couches de la formation de Niobrara, à l’exception des 2 supérieures ; en 2003, Kenneth Carpenter a étudié la distribution et la datation des fossiles de cette formation, démontrant que le Pteranodon sternbergi y existait il y a 88 à 85 millions d’années, tandis que le P. longiceps existait entre 86 et 84,5 millions d’années. Une troisième espèce possible, que Kellner a nommée le Geosternbergia maiseyi en 2010, est connue du membre Sharon Springs de la formation de schiste de pierre au Kansas, au Wyoming et au Dakota du Sud, datant d’il y a entre 81,5 et 80,5 millions d’années. Des fossiles de P. longiceps ont été trouvés dans des couches datant d’il y a 80 à 78,25 millions d’années.
Au début des années 1990, Bennett a noté que les 2 principales formes de ptéranodontes présentes dans la formation de Niobrara étaient précisément séparées dans le temps avec peu, voire aucun, chevauchement. Pour cette raison, et en raison de leur similitude globale grossière, il a suggéré qu’ils représentent probablement des chronoespèces au sein d’une seule lignée évolutive d’environ 4 millions d’années. En d’autres termes, une seule espèce de Pteranodon aurait été présente à un moment donné, et le P. sternbergi (ou Geosternbergia) était probablement l’espèce ancêtre directe du P. longiceps.
")
De nombreux chercheurs considèrent qu’il existe au moins 2 espèces de ptéranodon. Cependant, mis à part les différences entre les mâles et les femelles décrites ci-dessus, les squelettes post-crâniens de ptéranodon montrent peu ou pas de variation entre les espèces ou les spécimens, et le corps et les ailes de tous les ptéranodontes étaient essentiellement identiques.
2 espèces de ptéranodon sont traditionnellement reconnues comme valides : le Pteranodon longiceps, l’espèce-type, et le Pteranodon sternbergi. Les espèces ne diffèrent que par la forme de la crête chez les mâles adultes (décrite ci-dessus), et peut-être par l’angle de certains os du crâne. Parce que les fossiles de crânes de ptéranodon bien conservés sont extrêmement rares, les chercheurs utilisent la stratigraphie (c’est-à-dire la couche rocheuse de la formation géologique dans laquelle se trouve un fossile) pour déterminer l’identité des espèces dans la plupart des cas.
Le Pteranodon sternbergi est la seule espèce connue de Pteranodon avec une crête dressée. La mâchoire inférieure du P. sternbergi mesurait 1,25 mètre (4,1 pi) de long. Il a été collecté par George F. Sternberg en 1952 et décrit par John Christian Harksen en 1966, dans la partie inférieure de la formation de Niobrara. Il était plus ancien que le P. longiceps et est considéré par Bennett comme l’ancêtre direct de l’espèce ultérieure.
Étant donné que les fossiles identifiables comme le P. sternbergi se trouvent exclusivement dans les couches inférieures de la formation de Niobrara, et les fossiles de P. longiceps exclusivement dans les couches supérieures, un fossile dépourvu du crâne peut être identifié sur la base de sa position dans la colonne géologique (bien que pour de nombreuses découvertes de fossiles précoces, des données précises sur son emplacement n’aient pas été enregistrées, rendant de nombreux fossiles non identifiables).
|
En raison des variations subtiles entre les spécimens de ptéranodontidés de la formation de Niobrara, la plupart des chercheurs les ont tous attribués au seul genre Pteranodon, dans au moins 2 espèces (P. longiceps et P. sternbergi) qui se distinguent principalement par la forme de la crête. Cependant, la classification de ces 2 formes a varié d’un chercheur à l’autre. En 1972, Halsey Wilkinson Miller a publié un article soutenant que les différentes formes de Ptéranodon étaient suffisamment différentes pour être placées dans des sous-genres distincts. Il les nomma Ptéranodon (Occidentalia) occidentalis (pour l’espèce P. occidentalis, aujourd’hui désaffectée) et Ptéranodon (Sternbergia) sternbergi. Cependant, le nom Sternbergia était préoccupé, et en 1978, Miller a renommé l’espèce Pteranodon (Geosternbergia) sternbergi, et a nommé une troisième combinaison sous-genre/espèce pour le P. longiceps, comme Pteranodon (Longicepia) longiceps. Cependant, la plupart des chercheurs sur les ptérosaures de la fin du XXe siècle, y compris S. Christopher Bennett et Peter Wellnhofer, n’ont pas adopté ces noms sous-génériques et ont continué à placer toutes les espèces de ptéranodontes dans le seul genre Pteranodon.
En 2010, le chercheur sur les ptérosaures Alexander Kellner a revisité la classification de H.W. Miller. Kellner a suivi l’opinion de Miller selon laquelle les différences entre les espèces de Ptéranodon étaient suffisamment grandes pour les classer dans des genres différents. Il a placé le P. sternbergi dans le genre nommé par Miller, Geosternbergia, avec le spécimen de crâne des schistes de pierre que Bennett avait précédemment considéré comme un grand mâle P. longiceps. Kellner a fait valoir que la crête de ce spécimen, bien qu’incomplètement conservée, était la plus similaire au Geosternbergia. Parce que le spécimen était des millions d’années plus jeune que n’importe quel Geosternbergia connu, il l’a attribué à la nouvelle espèce Geosternbergia maiseyi. De nombreux autres spécimens de ptéranodontes sont connus de la même formation et de la même période, et Kellner a suggéré qu’ils pourraient appartenir à la même espèce que le G. maiseyi, mais parce qu’ils n’ont pas de crâne, il n’a pas pu les identifier avec certitude. Cependant, les 2 espèces précédemment désignées comme Geosternbergia ont été incluses séparément comme celles de Pteranodon (P. sternbergi et P. maiseyi) sur la base d’une analyse phylogénétique en 2024.

Un certain nombre d’autres espèces de ptéranodon ont été nommées depuis les années 1870, bien que la plupart soient maintenant considérées comme des synonymes juniors de 2 ou 3 espèces valides. La mieux soutenue est l’espèce type, P. longiceps, basée sur le spécimen bien conservé comprenant le premier crâne connu trouvé par S. W. Williston. Cet individu avait une envergure de 7 mètres (23 pieds). D’autres espèces valides incluent le P. sternbergi, peut-être plus grand, avec une envergure initialement estimée à 9 m (30 pi). Le P. oweni (P. occidentalis), le P. velox, le P. umbrosus, le P. harpyia et le P. comptus sont considérés comme des nomina dubia par Bennett (1994) et d’autres qui remettent en question leur validité. Tous sont probablement synonymes des espèces les plus connues.
Parce que la principale caractéristique distinctive que Marsh a notée pour le Ptéranodon était son manque de dents, tout fragment de mâchoire de ptérosaure édenté, où qu’il soit trouvé dans le monde, avait tendance à être attribué au Ptéranodon à la fin du XIXe et au début du XXe siècle. Il en a résulté une pléthore d’espèces et beaucoup de confusion. Le nom est devenu un taxon poubelle, un peu comme le dinosaure Megalosaurus, pour étiqueter tout reste de ptérosaure qui ne pouvait être distingué autrement que par l’absence de dents. Les espèces (souvent douteuses et dont on sait maintenant qu’elles sont basées sur la variation sexuelle ou les caractères juvéniles) ont été reclassifiées à plusieurs reprises, et plusieurs sous-genres ont été érigés dans les années 1970 par Halsey Wilkinson Miller pour les maintenir dans diverses combinaisons, ce qui a semé la confusion dans la taxonomie (les sous-genres comprennent Longicepia, Occidentalia et Geosternbergia). Parmi les auteurs notables qui ont discuté des différents aspects du ptéranodon, citons Bennett, Padian, Unwin, Kellner et Wellnhofer. 2 espèces, le P. oregonensis et le P. orientalis, ne sont pas des ptérodontidés et ont été renommées respectivement Bennettazhia oregonensis et Bogolubovia orientalis.
L’état des noms énumérés ci-dessous fait suite à une enquête menée par Bennett en 1994, sauf indication contraire.
| Nom | Auteur | Année | Status | Notes |
|---|---|---|---|---|
| Pterodactylus oweni | Marsh | 1871 | Nomen dubium | Renommé Pterodactylus occidentalis Marsh 1872 au motif qu'oweni était préoccupé par « Pterodactylus oweni » Seeley 1864 (nomen nudum pour Ornithocheirus oweni Seeley 1870) |
| Pterodactylus ingens | Marsh | 1872 | Reclassé en Pteranodon ingens | |
| Pterodactylus occidentalis | Marsh | 1872 | Objectif junior synonyme de Pterodactylus oweni | Reclassé de Pterodactylus oweni Marsh 1871 au motif que P. oweni était préoccupé par « Pterodactylus oweni » Seeley 1864 (nomen nudum pour Ornithocheirus oweni Seeley 1870) |
| Pterodactylus velox | Marsh | 1872 | Nomen dubium | Reclassé en Pteranodon velox |
| Ornithochirus umbrosus | Cope | 1872 | Nomen dubium | |
| Ornithochirus harpyia | Cope | 1872 | Nomen dubium | |
| Pterodactylus umbrosus | (Cope) Cope | (1872) 1874 | Reclassement de Ornithochirus umbrosus | |
| Pteranodon longiceps | Marsh | 1876 | Valide | Espèce type |
| Pteranodon ingens | (Marsh) Williston | (1872) 1876 | Nomen dubium | Reclassé de Pterodactylus ingens |
| Pteranodon occidentalis | Marsh | (1872) 1876 | Objectif junior synonyme de Pterodactylus oweni | Reclassé de Pterodactylus occidentalis |
| Pteranodon velox | Marsh | (1872) 1876 | Nomen dubium | Reclassé de Pterodactylus velox, basé sur un spécimen juvénile |
| Pteranodon gracilis | Marsh | 1876 | Reclassé en Nyctosaurus gracilis | |
| Pteranodon comptus | Marsh | 1876 | Nomen dubium | |
| Pteranodon nanus | Marsh | 1876 | Reclassé en Nyctosaurus nanus | |
| Ornithocheirus umbrosus | (Cope) Newton | (1872) 1888 | Reclassé en Pteranodon umbrosus | Correction orthographique d’Ornithochirus umbrosus |
| Ornithocheirus harpyia | (Cope) Newton | (1872) 1888 | Reclassé en Pteranodon harpyia | Correction orthographique d’Ornithochirus harpyia |
| Pteranodon umbrosus | (Cope) Williston | (1872) 1892 | Nomen dubium | Reclassement de Ornithochirus umbrosus |
| Ornithostoma ingens | (Marsh) Williston | (1872) 1893 | Synonyme de Pteranodon ingens | Reclassé de Pteranodon ingens |
| Ornithostoma umbrosum | (Cope) Williston | (1872) 1897 | Synonyme de Pteranodon umbrosus | Reclassé de Pteranodon umbrosus |
| Pteranodon oregonensis | Gilmore | 1928 | Reclassé en Bennettazhia oregonensis | |
| Pteranodon sternbergi | Harksen | 1966 | Valide | |
| Pteranodon marshi | Miller | 1972 | Synonyme de Pteranodon longiceps | |
| Pteranodon bonneri | Miller | 1972 | Reclassé en Nyctosaurus bonneri | |
| Pteranodon walkeri | Miller | 1972 | Synonyme de Pteranodon longiceps | |
| Pteranodon (Occidentalia) eatoni | (Miller) Miller | (1972) 1972 | Synonyme de Pteranodon sternbergi | |
| Pteranodon eatoni | (Miller) Miller | (1972) 1972 | Synonyme de Pteranodon sternbergi | Reclassé de Pteranodon (Occidentalia) eatoni |
| Pteranodon (Longicepia) longicps [sic] | (Marsh) Miller | (1872) 1972 | Synonyme de Pteranodon longiceps | Reclassé de Pteranodon longiceps |
| Pteranodon (Longicepia) marshi | (Miller) Miller | (1972) 1972 | Synonyme de Pteranodon longiceps | Reclassé de Pteranodon marshi |
| Pteranodon (Sternbergia) sternbergi | (Harksen) Miller | (1966) 1972 | Reclassé en Pteranodon (Geosternbergia) sternbergi | Reclassé de Pteranodon sternbergi |
| Pteranodon (Sternbergia) walkeri | (Miller) Miller | (1972) 1972 | Reclassé en Pteranodon (Geosternbergia) walkeri | Reclassé de Pteranodon walkeri |
| Pteranodon (Pteranodon) marshi | (Miller) Miller | (1972) 1973 | Synonyme de Pteranodon longiceps | Reclassé de Pteranodon marshi |
| Pteranodon (Occidentalia) occidentalis | (Marsh) Olshevsky | (1872) 1978 | Synonyme de Pteranodon occidentalis | Reclassé de Pteranodon occidentalis |
| Pteranodon (Longicepia) ingens | (Marsh) Olshevsky | (1872) 1978 | Synonyme de Pteranodon ingens | Reclassé de Pteranodon ingens |
| Pteranodon (Pteranodon) ingens | (Marsh) Olshevsky | (1872) 1978 | Synonyme de Pteranodon ingens | Reclassé de Pteranodon ingens |
| Pteranodon (Geosternbergia) walkeri | (Miller) Miller | (1972) 1978 | Synonyme de Pteranodon longiceps | Reclassé de Pteranodon walkeri |
| Pteranodon (Geosternbergia) sternbergi | (Harksen) Miller | (1966) 1978 | Synonyme de Pteranodon sternbergi | Reclassé de Pteranodon (Sternbergia) sternbergi |
| Pteranodon orientalis | (Bogolubov) Nesov & Yarkov | (1914) 1989 | Reclassé en Bogolubovia orientalis | Reclassé de Ornithostoma orientalis |
| Geosternbergia walkeri | (Miller) Olshevsky | (1972) 1991 | Synonyme de Pteranodon sternbergi | Reclassé de Pteranodon (Sternbergia) walkeri |
| Geosternbergia sternbergi | (Harksen) Olshevsky | (1966) 1991 | Synonyme de Pteranodon sternbergi | Reclassé de Pteranodon (Geosternbergia) sternbergi |






| 10 caractéristiques uniques qui vont vous épater ! par La encyclopédie des dinosaures | I'm a Pterodactyl - Dinosaur Songs par Howdytoons |
Rodan (1956) (空の大怪獣 ラドン, Sora no Daikaijū Radon) de Ishirō Honda
avec dans les rôles principaux :

|

|

|

|
|
(Shigeru Kawamura) |
(Kiyo) |
(Pr Kyōichirō Kashiwagi) |
(la jeune mariée) |
un ptéranodon mutant long d'une centaine de mètres et volant à la vitesse d'un avion hypersonique et appelé au Japon : kaiju Rodan.
Des mineurs sont attaqués dans un chantier près du mont Aso. Des policiers envoyés les chercher sont retrouvés déchiquetés. On identifie les coupables : des Meganulons, des larves de libellules préhistoriques géantes. Un des mineurs est retrouvé amnésique et en état de choc ; il déclare avoir vu un œuf géant éclore dans les mines. Quelque temps plus tard des avions sont détruits par un mystérieux objet volant.


Le kaiju Rodan apparaît dans 15 films, 5 séries/téléfilms, 17 jeux vidéos et 24 oeuvres de la littérature (livres, BD, manga, novel...)
Le Monde perdu : Jurassic Park (1997) (The Lost World: Jurassic Park) de Steven Spielberg
avec dans les rôles principaux :

|

|

|

|
|
(Pr Ian Malcolm) |
(Dr Sarah Harding) |
(John Hammond) |
(Kelly Curtis) |
un ptéranodon apparaît à la fin du film.
4 ans après le terrible fiasco de son Jurassic Park, le milliardaire John Hammond rappelle le Dr Ian Malcolm pour l'informer de son nouveau projet. Sur une île déserte, voisine du parc, vivent en liberté des centaines de dinosaures de toutes tailles et de toutes espèces. Ce sont des descendants des animaux clones en laboratoire. D'abord réticent, Ian se décide à rejoindre le docteur quand il apprend que sa fiancée fait partie de l'expédition scientifique. Il ignore qu'une autre expédition qui n'a pas les mêmes buts est également en route.


Jurassic Park 3 (2001) de Joe Johnston
avec dans les rôles principaux :

|

|

|

|
|
(Pr Alan Grant) |
(Pr Ellie Sattler) |
(Paul Kirby) |
(Amanda Kirby) |
les ptéranodons ont un rôle plus important, ils attaquent les héros du film dans une énorme volière. À la fin du film, ils arrivent à s’échapper de l’île et s'envolent pour trouver un nouvel endroit pour leurs nids.
Huit ans après la débâcle d´InGen, le paléontologue Alan Grant peine à trouver les moyens nécessaires pour continuer ses recherches. Grant accepte alors la proposition de l´aventurier Paul Kirby et de sa femme Amanda qui se disent prêts à financer ses travaux après qu´il les aura accompagnés dans leur tournée aérienne au-dessus du nouveau site de l´Ingen : Isla Sorna. Placée sous haute surveillance, cette île est devenue le principal centre d´élevage de dinosaures. Grant flaire l´entourloupe sitôt que le pilote amorce sa descente sur l´île.


Fidélité scientifique : Les ptéranodons de Jurassic Park 3, comme les vélociraptors, changent d'apparence par rapport à The Lost World sans explication officielle. Cette deuxième version est la plus souvent reprise par le contenu promotionnel, et est unanimement reconnue comme scientifiquement incorrecte pour un détail bien visible : les dents, ce qui contredit la signification de Pteranodon, « aile sans dent ».
L'animal entier a plus un physique de monstre de cinéma que de ptérosaure : la tête est trop petite – les ptérosaures ayant souvent des têtes disproportionnellement grosses par rapport à leur corps –, le bec est droit au lieu d'être incurvé et les 2 mâchoires sont de la même taille, alors que la supérieure devrait être légèrement plus longue. Le ptéra du film a une peau de cuir nue, et ne possède pas de couche de pycnofibres. Le bout de ses ailes est pointu au lieu d'arrondi, ses doigts sont trop gros et ses pieds sont préhensiles, comme des serres de rapaces, plutôt que plats et uniquement conçus pour la marche. Même s'il avait eu des serres, il aurait été incapable de soulever des humains, les ptérosaures étant des animaux légers et délicatement bâtis. Les cris stridents tiennent sans doute de la fiction : si on ignore le genre de sons que les ptérosaures produisaient, il est plus probable qu'ils émettaient des vocalisations semblables à celles des oiseaux ou des crocodiles. On suppose désormais que les ptérosaures ne nourrissaient pas leurs petits nichés sur des falaises comme les oiseaux, mais enterraient leurs œufs comme les tortues. Cela ne signifie pas qu'ils ne gardaient pas leur nid ou ne protégaient pas leurs petits. Les ptéras du film battent des ailes comme des chauve-souris et savent même voler latéralement. Ils ont été surdimensionnés, même si leur taille correspond aux estimations de l'époque.
Quelques bons côtés malgré tout : la longueur du cou, surestimée dans The Lost World, est corrigée, de même que la posture bipède : les ptéranodons sont correctement montrés comme des quadrupèdes, marchant sur leurs ailes repliées.
Jurassic World (2015) de Colin Trevorrow
avec dans les rôles principaux :

|

|

|

|
|
(Owen Grady) |
(Claire Dearing) |
(Vic Hoskins) |
(Barry Sembène) |
les ptéranodons. Dans une partie du film, les monstres s'attaquent à des visiteurs, en compagnie de dimorphodons.
8 ans après la débâcle d´InGen, le paléontologue Alan Grant peine à trouver les moyens nécessaires pour continuer ses recherches. Grant accepte alors la proposition de l´aventurier Paul Kirby et de sa femme Amanda qui se disent prêts à financer ses travaux après qu´il les aura accompagnés dans leur tournée aérienne au-dessus du nouveau site de l´Ingen : Isla Sorna. Placée sous haute surveillance, cette île est devenue le principal centre d´élevage de dinosaures. Grant flaire l´entourloupe sitôt que le pilote amorce sa descente sur l´île.


Fidélité scientifique : Troisième variante de Pteranodon de la franchise, elle est considérée comme étant la moins appréciée des 3. Il s'agit pourtant de la plus correcte : les principales différences sont l'absence de pycnofibres, le bout des ailes pointu plutôt qu'arrondi et les serres de rapaces préhensibles capables de soulever des humains et de s'agripper à des structures pour se poser, traits qu'elle partage avec les 2 premières versions. Le ptéranodon de Jurassic World présente d'autres divergences moins visibles, comme la taille et l'orientation des doigts, les arcades sourcillières trop épaisses, le corps trop maigre et la membrane des ailes qui descend jusqu'aux hanches et non jusqu'aux chevilles. Les ptéras du film sont aussi trop agressifs, et leur envergure, avoisinant 6 mètres, correspond à celle des mâles alors que ce sont toutes des femelles ; elles devraient donc être plus petites.
Jurassic World est cependant le premier film de la saga à montrer les femelles ayant des crêtes plus courtes. La forme du bec, légèrement incurvé vers le haut, est plus exacte que le bec droit des ptéras de Jurassic Park 3 ou le bec de cormoran de The Lost World. Le cou est plus court également, et le nouveau ptéra partage avec la variante du second film l'absence de dents, comme la signification de leur nom l'indique. Dans le film, les ptéranodons sont capables de plonger sous l'eau pour pêcher (ou dans le cas de Zara, repêcher) leurs proies. Il est très probable qu'ils en aient été capables dans la réalité, vivant à proximité des littoraux comme les oiseaux pêcheurs. Ils étaient donc les proies des prédateurs marins, ce que montre le film avec le mosasaure bondissant hors de l'eau pour en saisir un, possible clin d'œil à Fantasia.
Jurassic World: Fallen Kingdom (2018) de Juan Antonio Bayona
avec dans les rôles principaux :
|
|
|

|

|
|
(Owen Grady) |
(Claire Dearing) |
(Dr Zia Rodriguez) |
(Sir Benjamin Lockwood) |
des ptéranodons font partie des animaux capturés par les mercenaires, la scène post-générique du film montre trois d'entre eux survoler Las Vegas.
Cela fait maintenant 4 ans que les dinosaures se sont échappés de leurs enclos et ont détruit le parc à thème et complexe de luxe Jurassic World. Isla Nublar a été abandonnée par les humains alors que les dinosaures survivants sont livrés à eux-mêmes dans la jungle. Lorsque le volcan inactif de l'île commence à rugir, Owen et Claire s'organisent pour sauver les dinosaures restants de l'extinction. Owen se fait un devoir de retrouver Blue, son principal raptor qui a disparu dans la nature, alors que Claire, qui a maintenant un véritable respect pour ces créatures, s'en fait une mission...


Fidélité scientifique : Pour la première fois dans la franchise, les ptéranodons sont identiques à ceux du film précédent. En l'occurrence, ils sont toujours trop maigres, trop agressifs, manquent de pycnofibres, ont des ailes aux bouts pointus dont la membrane s'arrête aux hanches et des serres de rapace leur permettant de capturer des proies au sol et de se poser sur des structures.
Comme dans Jurassic World, ils ont des crêtes courtes de femelle et un bec édenté et incurvé comme dans la réalité. Auparavant massés dans une volière, ils sont à présent libres sur l'île et volent en groupes de plusieurs individus. On a retrouvé en Chine des fossiles de ptérosaures morts ensemble, ce qui indique qu'ils devaient vivre ensemble.
Nick Cutter et les Portes du temps (2007-2011) (Primeval) de Adrian Hodges et Tim Haines
avec dans les rôles principaux :

|

|

|

|
|
(Pr Nick Cutter) |
(Abigail « Abby » Sarah Maitland) |
(Stephen James Hart) |
(Connor Temple) |
un ptéranodon apparaît dans l'épisode 5 de la saison 1 (O.V.N.I.), il sème la panique sur un terrain de golf.
L'Angleterre est le théâtre d'étranges phénomènes : des créatures préhistoriques apparaissent aux quatre coins du Royaume... Le professeur Cutter, un homme brillant mais hanté par la disparition de sa femme, et son équipe, se lancent à leur poursuite et tentent de découvrir comment, ces espèces disparues depuis des milliers d'années, ont pu réapparaître soudainement...
Episode 5 : Un Ptéranodon apparaît sur un terrain de golf. Les ennuis commencent quand un golfeur est tué. Mais tout semble indiquer que ce n'est pas le Ptéranodon le coupable. Abby et Connor ne tardent pas à découvrir une nuée de ptérosaures carnivores tueurs attirés par le sang (peut-être des anurognathus).


Terra Nova (2011) de Kelly Marcel et Craig Silverstein
avec dans les rôles principaux :

|

|

|

|
|
(James « Jim » Shannon) |
(Dr Elisabeth Shannon) |
(Mira) |
(Skye « Godiche » Alexandria Tate) |
des ptéranodons survolent brièvement la jungle du Crétacé.
En l’an 2149, la vie sur Terre est menacée d'extinction par la pollution atmosphérique et les faibles ressources naturelles. La seule chance de sauver l’humanité est d’envoyer des individus à travers une faille spatio-temporelle, les menant 85 millions d'années en arrière vers un passé alternatif situé à la fin du Crétacé. La famille Shannon fait partie du dixième pèlerinage de colons pour Terra Nova, la première colonie humaine de l'autre côté du passage. Une nouvelle vie commence bientôt pour Jim, Elisabeth et leurs enfants Josh, Maddy et Zoé, malgré les dangers et le mystère des lieux...


Power Rangers : Dino Tonnerre (2004) (Power Rangers: Dino Thunder) de Eamon O'Sullivan
avec dans les rôles principaux :

|

|

|

|
|
(Conner McKnight (Ranger rouge)) |
(Ethan James (Ranger bleu)) |
(Kira Ford (Ranger jaune)) |
(Dr Thomas « Tommy » Oliver (Ranger noir)) |
le zord de la Ranger jaune est un ptéranodon
Le docteur Tommy Oliver, archéologue et ancien membre des Power Rangers, entreprend avec l'aide de son collègue Anton Mercer la création d'organismes à base d'ADN de dinosaure, destinés à aider à la protection de la planète. L'expérience tourne mal lorsque Mercer est transformé en une créature reptilienne du nom de Mesogog, et décide d'utiliser les créatures issues des recherches à son propre profit. Tommy réussit à faire exploser l'île où ont eu lieu les recherches, tuant apparemment Mesogog et ses forces, avant de fuir.
Quelque temps plus tard, alors que Tommy s'inscrit comme professeur dans un lycée, Mesogog resurgit de façon mystérieuse. Dans les incidents qui suivent, trois des élèves entrent en possession des Dinocristaux : Connor McKnight, un jeune footballeur immature, Ethan James, un passionné d'informatique et de jeux vidéo et Kira Ford, une jeune musicienne qui espère faire carrière. Ils obtiennent des pouvoirs et la capacité de contrôler les Dinozords créés par Tommy et Mercer. Tommy est alors contraint de les recruter et de les former en tant que nouveaux Power Rangers pour lutter contre Mesogog. Très peu de temps après, Tommy trouve un Dino Cristal noir, qui permet d'acquérir un pouvoir d'invisibilité et de rejoindre l'équipe de façon active.
Plus tard, un Ranger blanc les attaque. Il semble cependant reconnaitre Kira, qui parvient à le raisonner, et il décide de dévoiler son identité aux autres, qui n'en reviennent pas : il est en réalité Trent Mercer, le fils d'Anton, manipulé par Mesogog afin de les détruire. Trent leur avoue bien plus tard que Mesogog est en fait son père qui ne parvient pas à maîtriser ses transformations.


Dinotopia (2002) de Marco Brambilla
avec dans les rôles principaux :

|

|

|

|
|
(David Scott) |
(Karl Scott) |
(Marion Waldo) |
(Rosemary Waldo) |
des ptéranodons apparaissent dans l'épisode 1.
Naufragés, Karl et David échouent sur la terre inconnue de Dinotopia. Un monde où les humains co-habitent avec des dinosaures qui parlent. Une civilisation étrange et des rencontres étonnantes les attendent...


Le Dino Train (2009-2020) (Dinosaur Train) de Craig Bartlett
développé par Aqua System
La série de dessins animés Le Dino Train est construite autour de la famille Ptéranodons
Il raconte l'histoire de Samy, un jeune tyrannosaure adopté par une famille de ptéranodons, qui parcourt le monde des dinosaures à bord du Dino Train, pour découvrir les différentes espèces de dinosaures.


ARK: Survival Evolved (2017)
développé par Studio Wildcard, Instinct Games, Efecto Studios, Virtual Basement, édité par Studio Wildcard
il est possible d'apprivoiser un Ptéranodon
Dans le jeu, les joueurs doivent survivre dans un monde rempli de dinosaures, d'animaux préhistoriques et toutes sortes de créatures disparues, de dangers naturels et potentiellement de joueurs humains hostiles. Ils doivent aussi faire attention à leur vie, leur fatigue, leur poids, leur nourriture et leur soif, leur oxygène et leur torpeur ainsi que des dangers tels que les poisons, les maladies, la chaleur, le froid, etc. Le jeu peut être affiché en vue à la première ou troisième personne et les joueurs parcourent le monde ouvert et s'aventurent dans des endroits tels que les marécages, la toundra, la forêt de séquoia et ceci à pied, en chevauchant un dinosaure (ou autre créature montable) apprivoisé ou encore en volant. Pour cela il faudra au préalable fabriquer la selle spécifique du dinosaure apprivoisé.


Pokémon (1997-2025)
développé par Game Freak, édité par Nintendo, The Pokémon Company, distribué par Nintendo
un Ptéranodon. Le monstre Ptéra est inspiré d'un ptéranodon.
La série de jeux vidéo Pokémon est un ensemble de jeux vidéo de rôle fondé sur la franchise japonaise Pokémon. Le premier jeu sorti en 1997 est Pocket Monsters Vert et Rouge. Le dernier jeu qui est sorti en 2025 est Légendes Pokémon : Z-A.


Lost Eden (1995)
développé par Cryo Interactive, édité par Virgin Games, distribué par Rémi Herbulot
le personnage d’Eloi est un ptéranodon qui parle.
Le jeu se déroule dans un passé légendaire de la Terre où les humains et les dinosaures cohabitent et où les dinosaures ont une intelligence humaine. Le narrateur Eloi, un ptéranodon, nous conte a posteriori l'histoire du roi Adam de Mô, qu'il a connu dans sa jeunesse. Eloi jeune apparaît dans le jeu, où il fait partie des compagnons d'Adam, mais Eloi âgé fait quelques apparitions en tant que narrateur dans certaines scènes cinématiques.


Jurassic World Evolution (2018)
développé par Frontier Developments, édité par Frontier Developments et Universal Pictures, réalisé par Michael Brookes
un ptéranodon. Il est possible d'avoir des ptéranodons dans une volière.
Le joueur doit construire et gérer son parc à thème de dinosaure, comme dans le film Jurassic World. En plus de la gestion et la simulation, le jeu dispose aussi de nouvelles créatures. Le joueur pourra construire son parc sur l'île fictive d'Isla Nublar comme dans les films mais aussi sur d'autres îles. Le joueur devra également faire face à divers problèmes pouvant survenir à tout instant, comme des tempêtes, des coupures de courant, des évasions de dinosaures et même des cas d’espionnage industriel. Il peut même arriver de devoir faire face à plusieurs crises à la fois. Le joueur pourra aussi avoir la possibilité de prendre contrôle de certains véhicules durant les opérations.


One Piece (1997-présent) (ワンピース, Wan Pīsu) de Eiichirō Oda
un ptéranodon. La superstar King de l'équipage des cent bêtes a la capacité de se transformer en ptéranodon grâce à un fruit du démon.
L'histoire suit les aventures de Monkey D. Luffy, un garçon dont le corps a acquis les propriétés du caoutchouc après avoir mangé par inadvertance un fruit du démon. Avec son équipage de pirates, appelé l'équipage de Chapeau de paille, Luffy explore Grand Line à la recherche du trésor ultime connu sous le nom de « One Piece » afin de devenir le prochain roi des pirates.


un ptéranodon est sur un timbre postal de Géorgie en 1998.
L'histoire suit les aventures de Monkey D. Luffy, un garçon dont le corps a acquis les propriétés du caoutchouc après avoir mangé par inadvertance un fruit du démon. Avec son équipage de pirates, appelé l'équipage de Chapeau de paille, Luffy explore Grand Line à la recherche du trésor ultime connu sous le nom de « One Piece » afin de devenir le prochain roi des pirates.

Vous avez peut-être remarqué que je mets beaucoup plus de temps qu'avant pour publier la présentation d'une maquette. C'est simple : j'ai repris une de mes passions que j'avais abandonnée il y a 10 ans : la musique ou l'écriture et la composition de chansons.
Puis je me suis dit : pourquoi ne pas lier ma passion pour la musique avec celle pour la maquette et ce site Web ?
Ainsi, dorénavant et si c'est possible, j'écrirai une chanson pour le modèle que je présente et comme il faut un certain temps pour composer une chanson, j'utiliserai l'IA pour orchestrer la chanson.
Voici les chansons pour le Ptéranodon dont le titre est : L'Ombre de la Mer pour la chanson en français, The Winged and Toothless King pour la chanson en anglais, Rex Alatus Sine Dentibus pour la chanson en latin.
Comme le Ptéranodon est d'une époque où la musique n'existait pas, il m'a été très difficile de trouver quelque chose. Avec mon dernier album « Les Héros vol. 2 », j'ai parcouru des thèmes musicaux africain, oriental et amérindien, j'ai imaginé une orchestration Folk (guitare acoustique), percussions ethniques (tabla, tam-tam, darbuka, riqq), instruments arabe avec l'oud et moderne avec des synthétiseurs (les nappes). La chanson est interprétée par JJG (un des chanteurs de mon dernier album). Voici les paroles originales en français :
[Couplet 1]
Dans le ciel du Crétacé, quand le monde était vert,
Nul besoin d'atterrir sur le sol ou la terre.
Avec mes huit mètres d'envergure, je planais sans effort,
Vivant au-dessus des vagues, loin de tout le sort
Des lourds dinosaures qui marchaient sous les pins,
Mon royaume était l'azur, mes frères les dauphins.
Je suis le Ptéranodon, le maître des courants,
Portant mon ombre immense sur les sables mouvants.
[Couplet 2]
Le bec est incurvé, une mâchoire sans dent,
Pour attraper les poissons sur les flots du temps.
Mon crâne est décoré d'une crête osseuse en arrière,
Un long gouvernail étrange pour fendre l'atmosphère.
Elle m'aidait à voler, elle m'aidait à m'équilibrer,
Un signe de mon genre, un mâle pour me montrer.
J'étais léger comme une plume, un squelette creux,
Juste la force des muscles, l'instinct dans mes yeux.
[Refrain]
Je suis le Ptéranodon, la flèche qui s'élance,
L'aile faite de cuir, sans poils, sans méfiance.
Sur l'onde de l'Atlantique, je ne suis qu'une voile,
Une bête de l'air que rien, jamais, n'entrave ou n'avale.
Mon cri est celui du vent qui murmure le temps,
L'ombre qui survole l'océan, le géant planant.
[Couplet 3]
On me nomme « aile sans dent » (Pteranodon, c'est mon nom),
Je suis proche des dinos, mais ne suis point des leurs, non.
Je me lance à quatre pattes, c'est l'élan pour le ciel,
Je repousse la planète pour atteindre l'éternel.
Je mange du poisson, j'aime les crustacés,
La mer me nourrit, rien ne me fait stresser.
Mon habitat, c'est le Kansas, l'Amérique, c'est bien,
Mais mon cœur est partout où le ciel me retient.
[Refrain]
Je suis le Ptéranodon, la flèche qui s'élance,
L'aile faite de cuir, sans poils, sans méfiance.
Sur l'onde de l'Atlantique, je ne suis qu'une voile,
Une bête de l'air que rien, jamais, n'entrave ou n'avale.
Mon cri est celui du vent qui murmure le temps,
L'ombre qui survole l'océan, le géant planant.
[Pont]
Mais le jour est venu, le ciel est devenu l’enfer,
Quand le monde a tremblé sous un immense coup de tonnerre.
Le temps changea, la vie s'est éteinte, tout s'est rompu,
L'azur n'est plus mon ami, mon royaume est vaincu.
La fin de mon ère, la poussière et le froid,
Mais mon souvenir plane encore, juste après moi.
[Refrain]
Je suis le Ptéranodon, la flèche qui s'élance,
L'aile faite de cuir, sans poils, sans méfiance.
Sur l'onde de l'Atlantique, je ne suis qu'une voile,
Une bête de l'air que rien, jamais, n'entrave ou n'avale.
Mon cri est celui du vent qui murmure le temps,
L'ombre qui survole l'océan, le géant planant.
[Outro]
Le Ptéranodon... Juste un murmure d'aile dans la nuit de l’ère...
Un souvenir fossile... de l'air et de la mer...
La 2e chanson est en anglais sur un style de la chanson de Western avec un de mes chanteurs préférés : John M. (chanteur de The ballad of Athur Morgan (dans l'album « Nos Héros »), Le cri de la canopée, The Prophet's Brother (dans l'album « Nos Héros vol. 2 »)) :
[Verse 1]
In the late Cretaceous sky, a shadow starts to fly,
Not a feathered bird up high, just a glance as I pass by.
Over oceans I would glide, eight metres wide,
Let the wind my body guide, no place left to hide.
[Verse 2]
My bones were light and hollow, the way the current calls,
No teeth inside my swallow, watching for the squalls.
My long, sharp beak of bone, for the fishing that I'd do,
Over waves, all alone, hunting fish for my crew.
[Chorus]
I'm the Pteranodon, the Winged and Toothless King,
My crest is pointing back, the purpose that it brings.
A large, bony steering sail, cutting through the air,
We launch on all fours, an upward, sudden prayer,
To leave the rocky shore, for the soaring, sweet domain,
Never touching earth to walk, living in the rain.
[Verse 3]
We're kin to all the giants, the dinos on the ground,
But our lineage holds defiance, by the freedom that we found.
We lived near Western seas, where Kansas is today,
Flying with such ease, watching ages pass away.
[Chorus]
I'm the Pteranodon, the Winged and Toothless King,
My crest is pointing back, the purpose that it brings.
A large, bony steering sail, cutting through the air,
We launch on all fours, an upward, sudden prayer,
To leave the rocky shore, for the soaring, sweet domain,
Never touching earth to walk, living in the rain.
[Bridge]
The sky grew dark and cold, the mighty fire fell,
A story left untold, a final, fading spell.
The oceans ceased to call, the winds no longer blew,
We watched the empires fall, everything we knew.
[Chorus]
I'm the Pteranodon, the Winged and Toothless King,
My crest is pointing back, the purpose that it brings.
A large, bony steering sail, cutting through the air,
We launch on all fours, an upward, sudden prayer,
To leave the rocky shore, for the soaring, sweet domain,
Never touching earth to walk, living in the rain.
[Outro]
The long, long shadow...
Pteranodon longiceps... Just a memory in the stone.
La 3e chanson est toujours dans la langue de l'origine du modèle. Oui mais au crétacé, il n'y avait pas de langage humain. J'ai choisi le latin car c'est la langue des noms des animaux de cette époque. J'ai choisi un style de chant solennel avec très peu d'instruments : une contrebasse, un djembé et un violon. Voici les paroles :
[Verse 1]
In Caelo Cretaceo, umbra volare incipit,
Non avis plumosa, sed aura cito abit.
Super undas maris, octo metra late fuit,
Ventus corpus meum duxit, ubi nulla latebuit.
[Verse 2]
Ossa mea levia, sicut vocat fluentum,
Nulli dentes in ore, spectans ventum.
Rostrum longum et durum, pro piscatu meo,
Super fluctus solus, piscem capiens choro.
[Chorus]
Ego sum Pteranodon, Rex Alatus Sine Dentibus,
Crista mea retro monstrat, utitur pro omnibus.
Magna vela ossea, per aëra secans,
Quadrupes impellimur, precem altam petens,
Ut relinquamus litus, ad dominium dulce,
Numquam terram tangentes, viventes sub pluviae.
[Verse 3]
Nos ad magnos sumus affines, qui in terra ambulant,
Sed genus nostrum superbia per libertatem stat.
Prope maria occidentalia, ubi hodie Kansas iacet,
Volantes facilitate, ubi aetas transiit et tacet.
[Chorus]
Ego sum Pteranodon, Rex Alatus Sine Dentibus,
Crista mea retro monstrat, utitur pro omnibus.
Magna vela ossea, per aëra secans,
Quadrupes impellimur, precem altam petens,
Ut relinquamus litus, ad dominium dulce,
Numquam terram tangentes, viventes sub pluviae.
[Bridge]
Sed caelum frigidum est, ignis potens cecidit,
Fabula non dicta relictus est, somnium quod finidit.
Oceanus non vocat, ventus non amplius flat,
Regna ruentia vidimus, quod scivimus omne habitat.
[Chorus]
Ego sum Pteranodon, Rex Alatus Sine Dentibus,
Crista mea retro monstrat, utitur pro omnibus.
Magna vela ossea, per aëra secans,
Quadrupes impellimur, precem altam petens,
Ut relinquamus litus, ad dominium dulce,
Numquam terram tangentes, viventes sub pluviae.
[Outro]
Umbra longa...
Pteranodon longiceps...
Memoria in lapide.

Les dinosaures et les ptérosaures dominaient la Terre au cours de l'ère mésozoïque, et le ptéranodon ailé était un type de ptérosaure qui vivait vers la fin du Crétacé (il y a environ 85 millions d'années). Alors que son envergure était de 6 à 8 mètres, il ne pesait qu'environ 20 kilogrammes ; une membrane tendue depuis le quatrième doigt de leur main leur permettait de voler. Ils descendaient du ciel pour attraper leurs proies, parmi lesquelles on pense qu'il s'agissait de poissons.



|

|

|

|
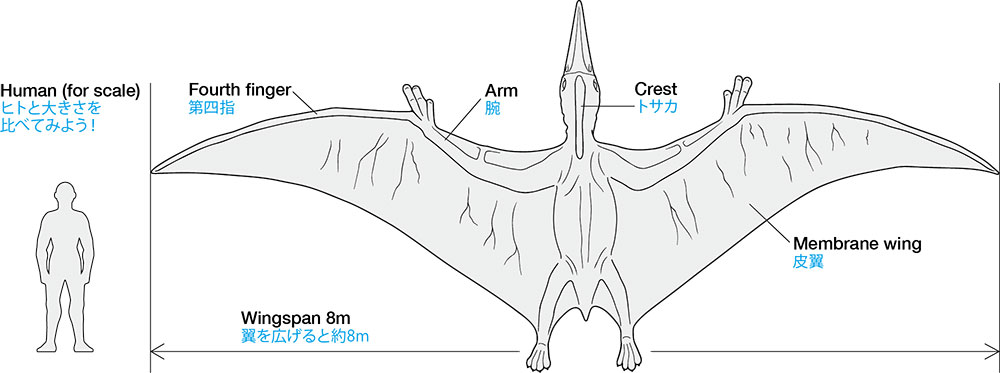
Vous pouvez le trouver pour l'instant sur :
 PlazaJapan à 3€59
PlazaJapan à 3€59
 El Taller del Modelista à 5€90
El Taller del Modelista à 5€90
 exito à 6€88
exito à 6€88
 WAM! Der Sockelshop à 6€93
WAM! Der Sockelshop à 6€93
 Dominomodel à 8€50
Dominomodel à 8€50
 OupsModel à 9€50
OupsModel à 9€50
Remarque : Les prix et la disponibilité ne sont donnés qu’à titre indicatif.
